
Growth Cones
Growth cones were recognised more than a hundred years ago by the famous spanish neuro-anatomist, Ramon y Cajal, as expansions at the tips of axons in fixed embryonic material. In 1910, Ross Harrison saw axons tipped with growth cones growing live across a microscopic slide from pieces of embyonic neural tube in tissue culture (Harrison, 1910).
Role of Growth Cones
Growth cones are highly motile structures that explore the extracellular environment, determine the direction of growth and then guide the extension of the axon in that direction. They assume several morphologies and may travel at different speeds as they navigate through parts of their pathways (Tosney and Landmesser, 1985; Bvolenta and Mason, 1987).
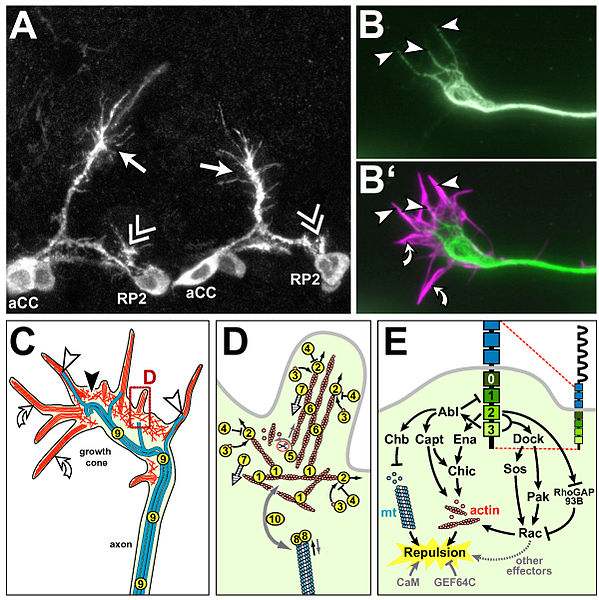
Image courtesy of https://commons.wikimedia.org/wiki/File:Drosophila_growth_cones_and_the_(potential)_factors_regulating_their_cytoskeletal_dynamics.jpg?uselang=en-gb
Growth cones that move along an established pathway,pioneered by other axons tend to be simple in shape whereas pioneer axons that are growing straight ahead have more traditional growth cones with several active filopodia and a few lamellipodia. Growth cones undergo dramatic changes when they arrive at choice points along the pathway. These changes in cell behaviour suggest that the growth cone has entered a novel environment and that now it must make choices based on its interaction with the environment. when it reaches its appropriate target, growth cones once again change their shape and growth rate.
Growing Zone
As growth cones move forward, they leave axons behind. This means that new material must be continually incorporated into the axon. In culture, when a particle is attached to a growing axon, they remain intact while the distal end of the axon elongates. This implies that membrane associated material is assembled at the distal end, the growth cone. The incorporation of this new material is calcium dependent, suggesting that the membrane is added by calcium mediated fusion of internal vesicles to the growth cone's surface. Supply of these vesicles seem to be from the central region of the growth cone, suggesting that majority of it is added at this point.
Addition of cytoskeletal components also take place primarily at the tip of the growing axon. Cytoskeletal components are not pushed forward , but assembled distally. In addition, pools of unassembled tubulin are concentrated in the growth cone which suggests that it is the most sensitive part of the axon to effects of microtubule depolymerizing agents such as nocodozole (Brown et al, 1992). Microtubule stabilizing proteins such as Tau are also highly concentrated near the growth cone, implying that is where unpolymerized tubulins are assembled into microtubules and stabilized.
From these studies, it is obvious that majority of axon growth and assembly of its parts takes place at the growth cone and an important function of growth cones is that of organizing the insertion of new component.